OK I am now officially completely driven insane by email. As part of my attempt to reduce email communication with people I am going to start posting some of the emails I get often into FAQs.
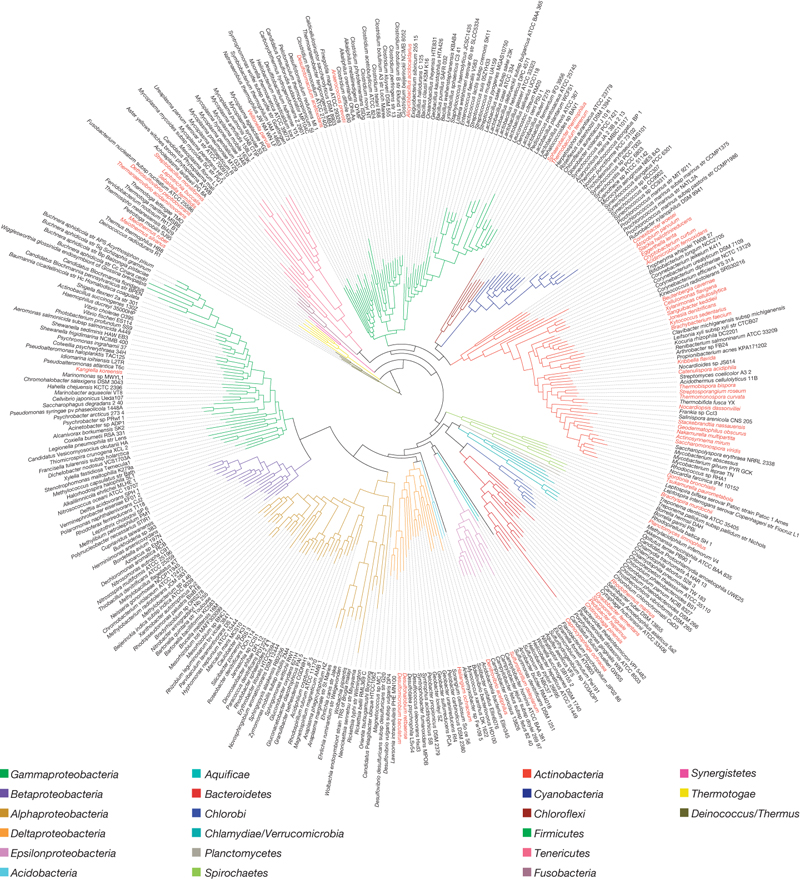
Today’s email relates to the 2009 paper on a “Phylogeny driven genomic encyclopedia of bacteria and archaea” for which I was the senior and corresponding author. The email is asking for higher resolution figures that were published in the paper. This person and many others have asked for a higher res. version of our “genome tree” which was Figure 1. Here is the version from the paper
But alas, as a JPG when you zoom in you can’t see the text very well. And about 30 or so people, maybe more, have asked for a higher res. version. Well, the simplest way to get this figure with legible fonts when zoomed in is to get the PDF of the paper and zoom in on it. But that may not be for everyone – so here is a link to the PDF of the figure that I posted on postures (Blogger does not allow PDF uploads). I also posted PDFs of the other figures.
Many people also ask for the treefile (which is basically a coded version of the phylogenetic tree for viewing and analysis). I am directly posting the treefile below and have also submitted it to “Treebase” (which we should have done before). Enjoy … and in the future I will be pointing people to this page when they ask for the figure/treefile. Not sure this will have saved me anytime but am sick of writing a lot of this in emails back to people …
#NEXUS
BEGIN trees;
TREE ‘Tree1’ = (((((‘GEBA_Thermanaerovibrio_acidaminovorans’:0.190689,’GEBA_Dethiosulfovibrio_peptidovorans’:0.263143):0.276658,((((((((((((((((((((((((((((((((((((((((‘Escherichia_coli_O157_H7_str_Sakai’:0.0,’Escherichia_coli_str_K12_substr_MG1655′:0.0):1.51E-4,’Escherichia_coli_str_K12_substr_DH10B’:0.0):2.0E-5,(‘Escherichia_coli_ATCC_8739′:1.7E-4,’Escherichia_coli_HS’:1.71E-4):1.7E-4):0.0,(‘Escherichia_coli_536′:0.001196,’Escherichia_coli_APEC_O1’:0.0):0.0):0.0,((‘Shigella_flexneri_2a_str_301′:0.001883,’Shigella_flexneri_2a_str_2457T’:0.0):1.71E-4,’Shigella_flexneri_5_str_8401′:3.44E-4):5.12E-4):0.0,(‘Shigella_boydii_CDC_3083_94′:5.13E-4,’Shigella_boydii_Sb227′:8.57E-4):3.42E-4):0.0,’Escherichia_coli_SMS_3_5′:0.0):0.0,’Escherichia_coli_O157_H7_EDL933′:0.0):0.0,’Escherichia_coli_CFT073′:0.0):0.0,’Escherichia_coli_UTI89′:0.0):0.0,’Escherichia_coli_E24377A’:0.0):0.0,’Shigella_dysenteriae_Sd197′:0.001025):1.69E-4,’Shigella_sonnei_Ss046′:0.001028):0.012167,(((((((‘Salmonella_enterica_subsp_enterica_serovar_Typhi_str_Ty2′:0.0,’Salmonella_enterica_subsp_enterica_serovar_Typhi_str_CT18′:1.7E-4):3.4E-4,’Salmonella_typhimurium_LT2′:0.0):0.0,’Salmonella_enterica_subsp_enterica_serovar_Paratyphi_A_str_ATCC_9150′:8.5E-4):2.0E-6,’Salmonella_enterica_subsp_enterica_serovar_Paratyphi_B_str_SPB7′:1.8E-4):1.69E-4,’Salmonella_enterica_subsp_enterica_serovar_Choleraesuis_str_SC_B67′:3.48E-4):0.001774,’Salmonella_enterica_subsp_arizonae_serovar_62_z4_z23_’:0.001673):0.008533,’Citrobacter_koseri_ATCC_BAA_895′:0.007106):0.003103):0.00437,’Klebsiella_pneumoniae_subsp_pneumoniae_MGH_78578′:0.012094):0.004392,’Enterobacter_sakazakii_ATCC_BAA_894′:0.019781):0.007021,’Enterobacter_sp_638′:0.019532):0.027075,(‘Erwinia_tasmaniensis’:0.033979,’Serratia_proteamaculans_568′:0.033607):0.013319):0.008604,’Erwinia_carotovora_subsp_atroseptica_SCRI1043′:0.03122):0.012206,((((((‘Yersinia_pestis_KIM’:0.0,’Yersinia_pseudotuberculosis_IP_31758′:0.0):0.0,’Yersinia_pestis_CO92′:0.0):0.0,’Yersinia_pseudotuberculosis_IP_32953′:0.0):0.0,((((‘Yersinia_pseudotuberculosis_PB1_’:0.0,’Yersinia_pestis_Pestoides_F’:0.0):0.0,(‘Yersinia_pseudotuberculosis_YPIII’:0.0,’Yersinia_pestis_Nepal516′:0.0):0.0):0.0,’Yersinia_pestis_Antiqua’:1.72E-4):0.0,’Yersinia_pestis_biovar_Microtus_str_91001′:0.0):0.0):0.0,’Yersinia_pestis_Angola’:3.44E-4):0.00575,’Yersinia_enterocolitica_subsp_enterocolitica_8081′:0.009468):0.030284):0.008222,((((‘Candidatus_Blochmannia_pennsylvanicus_str_BPEN’:0.111832,’Candidatus_Blochmannia_floridanus’:0.199319):0.14508,(((‘Buchnera_aphidicola_str_APS_Acyrthosiphon_pisum_’:0.081646,’Buchnera_aphidicola_str_Sg_Schizaphis_graminum_’:0.078489):0.090786,’Buchnera_aphidicola_str_Bp_Baizongia_pistaciae_’:0.232468):0.043831,(‘Wigglesworthia_glossinidia_endosymbiont_of_Glossina_brevipalpis’:0.30099,’Buchnera_aphidicola_str_Cc_Cinara_cedri_’:0.303142):0.063766):0.064196):0.054991,’Baumannia_cicadellinicola_str_Hc_Homalodisca_coagulata_’:0.166451):0.081445,’Sodalis_glossinidius_str_morsitans_’:0.026696):0.024889):0.01239,’Photorhabdus_luminescens_subsp_laumondii_TTO1′:0.048195):0.050763,((((‘Haemophilus_somnus_129PT’:9.73E-4,’Haemophilus_somnus_2336′:7.28E-4):0.038446,’Pasteurella_multocida_subsp_multocida_str_Pm70′:0.033371):0.011204,((‘Mannheimia_succiniciproducens_MBEL55E’:0.02899,’Actinobacillus_succinogenes_130Z’:0.035825):0.013874,(((‘Haemophilus_influenzae_Rd_KW20′:0.002043,’Haemophilus_influenzae_PittGG’:0.00105):5.49E-4,’Haemophilus_influenzae_86_028NP’:5.15E-4):2.24E-4,’Haemophilus_influenzae_PittEE’:0.001026):0.040304):0.008478):0.011217,(((‘Actinobacillus_pleuropneumoniae_L20′:0.00137,’Actinobacillus_pleuropneumoniae_serovar_7_str_AP76′:3.39E-4):1.74E-4,’Actinobacillus_pleuropneumoniae_serovar_3_str_JL03′:0.003062):0.011765,’Haemophilus_ducreyi_35000HP’:0.024577):0.038512):0.080775):0.047571,(((((‘Vibrio_cholerae_O1_biovar_eltor_str_N16961′:3.64E-4,’Vibrio_cholerae_O395’:8.38E-4):0.041305,(‘Vibrio_vulnificus_CMCP6′:5.27E-4,’Vibrio_vulnificus_YJ016’:1.72E-4):0.020829):0.011445,(‘Vibrio_parahaemolyticus_RIMD_2210633′:0.00625,’Vibrio_harveyi_ATCC_BAA_1116′:0.010839):0.011481):0.027253,’Vibrio_fischeri_ES114′:0.05376):0.027779,’Photobacterium_profundum_SS9’:0.072105):0.054993):0.023142,(‘Aeromonas_hydrophila_subsp_hydrophila_ATCC_7966′:0.006284,’Aeromonas_salmonicida_subsp_salmonicida_A449’:0.0108):0.109795):0.020707,((((((‘Shewanella_halifaxensis_HAW_EB4′:0.008905,’Shewanella_pealeana_ATCC_700345’:0.003805):0.025082,(‘Shewanella_sediminis_HAW_EB3′:0.019306,’Shewanella_woodyi_ATCC_51908′:0.012326):0.017175):0.016987,’Shewanella_loihica_PV_4’:0.021173):0.024752,((((((‘Shewanella_sp_ANA_3′:0.002092,’Shewanella_sp_MR_4′:9.8E-4):7.54E-4,’Shewanella_sp_MR_7′:3.48E-4):0.003036,’Shewanella_oneidensis_MR_1’:0.002265):0.010027,(‘Shewanella_sp_W3_18_1′:3.26E-4,’Shewanella_putrefaciens_CN_32’:1.85E-4):0.001996):0.006463,((‘Shewanella_baltica_OS155′:0.001236,’Shewanella_baltica_OS195′:0.0):5.23E-4,’Shewanella_baltica_OS185’:6.84E-4):0.009966):0.015626,(‘Shewanella_denitrificans_OS217′:0.017657,’Shewanella_frigidimarina_NCIMB_400′:0.02512):0.022711):0.020469):0.018183,’Shewanella_amazonensis_SB2B’:0.035265):0.092511,’Psychromonas_ingrahamii_37′:0.170059):0.021406):0.025447,((‘Idiomarina_loihiensis_L2TR’:0.129215,’Pseudoalteromonas_atlantica_T6c’:0.121204):0.026755,(‘Pseudoalteromonas_haloplanktis_TAC125′:0.120366,’Colwellia_psychrerythraea_34H’:0.144993):0.031198):0.015669):0.082415,’GEBA_Kangiella_koreensis’:0.172853):0.0486,((((((‘Acinetobacter_baumannii_ACICU’:8.84E-4,’Acinetobacter_baumannii_ATCC_17978′:0.003413):5.27E-4,(‘Acinetobacter_baumannii_SDF’:8.48E-4,’Acinetobacter_baumannii_AYE’:1.7E-4):0.0):0.020056,’Acinetobacter_sp_ADP1′:0.027905):0.107102,((‘Psychrobacter_arcticus_273_4′:0.006888,’Psychrobacter_cryohalolentis_K5′:0.006086):0.066403,’Psychrobacter_sp_PRwf_1′:0.060503):0.121017):0.15381,’Alcanivorax_borkumensis_SK2’:0.175254):0.040455,(((‘Chromohalobacter_salexigens_DSM_3043′:0.167612,’Marinomonas_sp_MWYL1’:0.175719):0.038094,(‘Marinobacter_aquaeolei_VT8′:0.1125,’Hahella_chejuensis_KCTC_2396’:0.130701):0.053467):0.019075,(((((((((‘Pseudomonas_syringae_pv_syringae_B728a’:0.001801,’Pseudomonas_syringae_pv_phaseolicola_1448A’:0.002321):0.002597,’Pseudomonas_syringae_pv_tomato_str_DC3000′:0.003918):0.02921,’Pseudomonas_fluorescens_Pf_5′:0.014609):0.00751,’Pseudomonas_fluorescens_PfO_1′:0.010753):0.026949,((((‘Pseudomonas_putida_KT2440′:1.71E-4,’Pseudomonas_putida_F1′:0.0):0.001622,’Pseudomonas_putida_GB_1′:0.003034):0.003248,’Pseudomonas_putida_W619′:0.007868):0.003675,’Pseudomonas_entomophila_L48′:0.007306):0.02498):0.030505,’Pseudomonas_mendocina_ymp’:0.030084):0.018411,’Pseudomonas_stutzeri_A1501′:0.035067):0.021645,((‘Pseudomonas_aeruginosa_PAO1′:0.0,’Pseudomonas_aeruginosa_UCBPP_PA14′:1.71E-4):0.001113,’Pseudomonas_aeruginosa_PA7’:0.001132):0.032184):0.118537,(‘Saccharophagus_degradans_2_40′:0.107494,’Cellvibrio_japonicus_Ueda107’:0.109149):0.090952):0.024456):0.029903):0.041423):0.029921,(((‘Legionella_pneumophila_str_Corby’:0.001506,’Legionella_pneumophila_subsp_pneumophila_str_Philadelphia_1′:5.43E-4):3.1E-4,(‘Legionella_pneumophila_str_Lens’:0.00222,’Legionella_pneumophila_str_Paris’:0.001831):3.76E-4):0.255271,((‘Coxiella_burnetii_RSA_331′:0.00102,’Coxiella_burnetii_RSA_493′:5.1E-4):0.001141,’Coxiella_burnetii_Dugway_5J108_111’:0.001413):0.286937):0.063734):0.020357,((((((((‘Francisella_tularensis_subsp_tularensis_SCHU_S4′:0.0,’Francisella_tularensis_subsp_tularensis_FSC198′:0.0):0.001507,’Francisella_tularensis_subsp_tularensis_WY96_3418’:3.35E-4):3.34E-4,((‘Francisella_tularensis_subsp_holarctica_OSU18′:0.0,’Francisella_tularensis_subsp_holarctica’:5.02E-4):0.0,’Francisella_tularensis_subsp_holarctica_FTNF002_00′:3.35E-4):0.002852):0.0,’Francisella_tularensis_subsp_mediasiatica_FSC147′:0.002516):0.001333,’Francisella_tularensis_subsp_novicida_U112′:6.81E-4):0.02679,’Francisella_philomiragia_subsp_philomiragia_ATCC_25017′:0.028537):0.343835,’Dichelobacter_nodosus_VCS1703A’:0.35117):0.058577,((‘Candidatus_Ruthia_magnifica_str_Cm_Calyptogena_magnifica_’:0.048117,’Candidatus_Vesicomyosocius_okutanii_HA’:0.070088):0.299069,’Thiomicrospira_crunogena_XCL_2′:0.218606):0.069384):0.027865):0.019953,(((‘Halorhodospira_halophila_SL1′:0.175731,’Alkalilimnicola_ehrlichei_MLHE_1′:0.10505):0.103584,’Nitrosococcus_oceani_ATCC_19707’:0.225193):0.031953,((((((‘Xanthomonas_campestris_pv_campestris_str_8004′:3.43E-4,’Xanthomonas_campestris_pv_campestris_str_ATCC_33913′:5.15E-4):3.45E-4,’Xanthomonas_campestris_pv_campestris’:1.7E-4):0.006192,(((‘Xanthomonas_oryzae_pv_oryzae_KACC10331′:1.75E-4,’Xanthomonas_oryzae_pv_oryzae_MAFF_311018′:0.0):0.0,’Xanthomonas_oryzae_pv_oryzae_PXO99A’:0.002957):0.008947,(‘Xanthomonas_campestris_pv_vesicatoria_str_85_10′:0.001996,’Xanthomonas_axonopodis_pv_citri_str_306′:0.00127):0.004934):0.005782):0.031343,’Stenotrophomonas_maltophilia_K279a’:0.060056):0.030521,(((‘Xylella_fastidiosa_M23′:0.0,’Xylella_fastidiosa_Temecula1′:1.74E-4):0.005446,’Xylella_fastidiosa_M12′:0.003419):0.005622,’Xylella_fastidiosa_9a5c’:0.003688):0.09616):0.251782,’Methylococcus_capsulatus_str_Bath’:0.208011):0.028921):0.02517):0.056558,((((((‘Nitrosomonas_europaea_ATCC_19718′:0.04084,’Nitrosomonas_eutropha_C91′:0.04959):0.144982,’Nitrosospira_multiformis_ATCC_25196′:0.08516):0.076439,’Thiobacillus_denitrificans_ATCC_25259′:0.145726):0.023371,’Methylobacillus_flagellatus_KT’:0.155554):0.027233,(((‘Azoarcus_sp_BH72′:0.052172,’Azoarcus_sp_EbN1′:0.058776):0.062917,’Dechloromonas_aromatica_RCB’:0.113139):0.05407,((((((‘Polynucleobacter_sp_QLW_P1DMWA_1′:0.011,’Polynucleobacter_necessarius_STIR1’:0.023235):0.136644,((((‘Cupriavidus_taiwanensis’:0.005601,’Ralstonia_eutropha_H16′:0.005264):0.006279,’Ralstonia_eutropha_JMP134′:0.007605):0.010799,’Ralstonia_metallidurans_CH34′:0.013934):0.029247,(‘Ralstonia_solanacearum_GMI1000′:0.02203,’Ralstonia_pickettii_12J’:0.021784):0.025195):0.037629):0.022794,(((‘Burkholderia_phytofirmans_PsJN’:0.004098,’Burkholderia_xenovorans_LB400′:0.002629):0.017129,’Burkholderia_phymatum_STM815′:0.017185):0.015186,(((((‘Burkholderia_ambifaria_AMMD’:0.001201,’Burkholderia_ambifaria_MC40_6′:1.65E-4):0.00415,(((‘Burkholderia_cenocepacia_AU_1054′:0.0,’Burkholderia_cenocepacia_HI2424′:1.71E-4):1.71E-4,’Burkholderia_cenocepacia_MC0_3′:0.0):0.002317,’Burkholderia_sp_383′:0.007333):0.002889):0.00261,’Burkholderia_vietnamiensis_G4′:0.003346):0.015421,’Burkholderia_multivorans_ATCC_17616’:0.005208):0.005853,((((((((‘Burkholderia_mallei_NCTC_10247′:0.0,’Burkholderia_mallei_NCTC_10229′:1.72E-4):3.43E-4,’Burkholderia_mallei_ATCC_23344′:1.91E-4):0.0,’Burkholderia_mallei_SAVP1′:0.0):1.71E-4,’Burkholderia_pseudomallei_K96243′:5.16E-4):0.0,’Burkholderia_pseudomallei_1710b’:0.0):0.0,’Burkholderia_pseudomallei_668′:0.0):0.0,’Burkholderia_pseudomallei_1106a’:1.71E-4):0.004036,’Burkholderia_thailandensis_E264′:0.005011):0.007936):0.019911):0.063072):0.026718,(‘Herminiimonas_arsenicoxydans’:0.021781,’Janthinobacterium_sp_Marseille’:0.013485):0.104015):0.024622,((‘Methylibium_petroleiphilum_PM1′:0.071447,’Leptothrix_cholodnii_SP_6’:0.085431):0.041536,((((‘Delftia_acidovorans_SPH_1′:0.061373,’Acidovorax_sp_JS42′:0.025396):0.016116,’Acidovorax_avenae_subsp_citrulli_AAC00_1′:0.025052):0.01365,’Verminephrobacter_eiseniae_EF01_2’:0.072504):0.026641,((‘Polaromonas_naphthalenivorans_CJ2′:0.031581,’Polaromonas_sp_JS666′:0.022265):0.044515,’Rhodoferax_ferrireducens_T118’:0.06856):0.022175):0.065841):0.119404):0.025568,((((‘Bordetella_bronchiseptica_RB50′:1.72E-4,’Bordetella_parapertussis_12822′:0.001377):1.18E-4,’Bordetella_pertussis_Tohama_I’:0.001433):0.021534,’Bordetella_avium_197N’:0.031614):0.010355,’Bordetella_petrii_DSM_12804′:0.021324):0.132685):0.071258):0.027059):0.037078,(((‘Neisseria_gonorrhoeae_FA_1090′:5.0E-4,’Neisseria_gonorrhoeae_NCCP11945’:0.001485):0.006103,(((‘Neisseria_meningitidis_MC58′:0.001366,’Neisseria_meningitidis_Z2491′:0.00101):7.87E-4,’Neisseria_meningitidis_FAM18′:0.001575):5.57E-4,’Neisseria_meningitidis_053442′:0.002602):0.001738):0.167934,’Chromobacterium_violaceum_ATCC_12472’:0.09855):0.087498):0.124859):0.205339,(((((((((((((‘Rhizobium_etli_CIAT_652′:0.003437,’Rhizobium_etli_CFN_42′:0.005094):0.006043,’Rhizobium_leguminosarum_bv_viciae_3841′:0.011645):0.036083,’Agrobacterium_tumefaciens_str_C58’:0.045233):0.016839,(‘Sinorhizobium_meliloti_1021′:0.006124,’Sinorhizobium_medicae_WSM419’:0.006258):0.029756):0.070623,(((((‘Bartonella_henselae_str_Houston_1′:0.020444,’Bartonella_quintana_str_Toulouse’:0.029388):0.009808,’Bartonella_tribocorum_CIP_105476′:0.025338):0.031387,’Bartonella_bacilliformis_KC583′:0.065286):0.098029,(((((‘Brucella_canis_ATCC_23365′:6.88E-4,’Brucella_suis_1330′:1.72E-4):6.89E-4,’Brucella_suis_ATCC_23445’:0.001033):0.0,(((‘Brucella_abortus_S19′:1.76E-4,’Brucella_melitensis_biovar_Abortus_2308′:1.72E-4):0.0,’Brucella_abortus_biovar_1_str_9_941′:0.0):6.84E-4,’Brucella_melitensis_16M’:0.002611):1.71E-4):0.0,’Brucella_ovis_ATCC_25840′:0.001206):0.013975,’Ochrobactrum_anthropi_ATCC_49188′:0.01275):0.047742):0.034988,(‘Mesorhizobium_loti_MAFF303099′:0.077568,’Mesorhizobium_sp_BNC1’:0.07419):0.027327):0.021349):0.086016,((((((‘Methylobacterium_extorquens_PA1′:0.011368,’Methylobacterium_populi_BJ001′:0.004799):0.03256,’Methylobacterium_radiotolerans_JCM_2831′:0.045067):0.041026,’Methylobacterium_sp_4_46′:0.058056):0.074338,’Beijerinckia_indica_subsp_indica_ATCC_9039’:0.149061):0.026115,(‘Xanthobacter_autotrophicus_Py2′:0.058512,’Azorhizobium_caulinodans_ORS_571’:0.044976):0.079042):0.02072,((((‘Bradyrhizobium_sp_ORS278′:0.007711,’Bradyrhizobium_sp_BTAi1′:0.006207):0.029183,’Bradyrhizobium_japonicum_USDA_110’:0.033032):0.020122,(((‘Rhodopseudomonas_palustris_TIE_1′:0.0,’Rhodopseudomonas_palustris_CGA009’:3.43E-4):0.030733,(‘Rhodopseudomonas_palustris_BisB5′:0.012815,’Rhodopseudomonas_palustris_HaA2’:0.011805):0.010984):0.018577,(‘Rhodopseudomonas_palustris_BisB18′:0.037297,’Rhodopseudomonas_palustris_BisA53’:0.031869):0.014739):0.023424):0.01122,(‘Nitrobacter_winogradskyi_Nb_255′:0.018532,’Nitrobacter_hamburgensis_X14′:0.017835):0.041031):0.111343):0.056949):0.041219,’Parvibaculum_lavamentivorans_DS_1’:0.17312):0.029751,((‘Maricaulis_maris_MCS10′:0.176054,’Hyphomonas_neptunium_ATCC_15444’:0.253873):0.037937,(‘Caulobacter_sp_K31′:0.046364,’Caulobacter_crescentus_CB15’:0.03404):0.177012):0.067981):0.024054,(((((‘Silicibacter_pomeroyi_DSS_3′:0.054186,’Silicibacter_sp_TM1040′:0.048369):0.026318,’Roseobacter_denitrificans_OCh_114′:0.076151):0.022514,’Jannaschia_sp_CCS1′:0.099178):0.015502,’Dinoroseobacter_shibae_DFL_12’:0.062324):0.048422,(((‘Rhodobacter_sphaeroides_ATCC_17029′:3.54E-4,’Rhodobacter_sphaeroides_2_4_1′:7.92E-4):0.0133,’Rhodobacter_sphaeroides_ATCC_17025′:0.013986):0.076691,’Paracoccus_denitrificans_PD1222’:0.091793):0.030158):0.202704):0.030795,(((‘Novosphingobium_aromaticivorans_DSM_12444′:0.086704,’Erythrobacter_litoralis_HTCC2594′:0.121244):0.045873,’Sphingopyxis_alaskensis_RB2256’:0.085216):0.03517,(‘Sphingomonas_wittichii_RW1′:0.074921,’Zymomonas_mobilis_subsp_mobilis_ZM4’:0.113715):0.03071):0.187787):0.027917,((‘Rhodospirillum_rubrum_ATCC_11170′:0.169238,’Magnetospirillum_magneticum_AMB_1’:0.125062):0.055149,(((‘Gluconobacter_oxydans_621H’:0.101123,’Gluconacetobacter_diazotrophicus_PAl_5′:0.059111):0.060663,’Granulibacter_bethesdensis_CGDNIH1′:0.085514):0.032014,’Acidiphilium_cryptum_JF_5′:0.120348):0.145099):0.055072):0.09899,(((((‘Rickettsia_bellii_RML369_C’:6.4E-4,’Rickettsia_bellii_OSU_85_389′:6.24E-4):0.053585,(((‘Rickettsia_typhi_str_Wilmington’:0.015422,’Rickettsia_prowazekii_str_Madrid_E’:0.012042):0.039247,((‘Rickettsia_felis_URRWXCal2′:0.00684,’Rickettsia_akari_str_Hartford’:0.024586):0.00502,(((‘Rickettsia_rickettsii_str_Iowa’:0.0,’Rickettsia_rickettsii_str_Sheila_Smith_’:0.0):0.00663,’Rickettsia_conorii_str_Malish_7′:0.002765):0.00386,’Rickettsia_massiliae_MTU5′:0.00794):0.008218):0.002179):0.00446,’Rickettsia_canadensis_str_McKiel’:0.030982):0.03239):0.199956,(‘Orientia_tsutsugamushi_Boryong’:0.012903,’Orientia_tsutsugamushi_str_Ikeda’:0.003049):0.386268):0.180586,((((‘Anaplasma_phagocytophilum_HZ’:0.123173,’Anaplasma_marginale_str_St_Maries’:0.120008):0.159306,((‘Ehrlichia_ruminantium_str_Gardel’:0.002292,’Ehrlichia_ruminantium_str_Welgevonden’:0.002453):0.053871,(‘Ehrlichia_canis_str_Jake’:0.027749,’Ehrlichia_chaffeensis_str_Arkansas’:0.027236):0.031378):0.134774):0.151991,((‘Wolbachia_pipientis’:0.072061,’Wolbachia_endosymbiont_of_Drosophila_melanogaster’:0.04265):0.018425,’Wolbachia_endosymbiont_strain_TRS_of_Brugia_malayi’:0.077353):0.269684):0.141145,’Neorickettsia_sennetsu_str_Miyayama’:0.697366):0.171539):0.064736,’Candidatus_Pelagibacter_ubique_HTCC1062′:0.549831):0.056614):0.089191,’Magnetococcus_sp_MC_1′:0.339568):0.063213):0.093791,((‘Acidobacteria_bacterium_Ellin345′:0.221486,’Solibacter_usitatus_Ellin6076’:0.235193):0.269435,((((((‘Geobacter_sulfurreducens_PCA’:0.05188,’Geobacter_metallireducens_GS_15′:0.04331):0.045927,’Geobacter_uraniireducens_Rf4′:0.08407):0.035503,(‘Pelobacter_propionicus_DSM_2379′:0.107415,’Geobacter_lovleyi_SZ’:0.104313):0.062144):0.11137,’Pelobacter_carbinolicus_DSM_2380′:0.218315):0.102231,(((((((‘Desulfovibrio_vulgaris_subsp_vulgaris_DP4′:2.75E-4,’Desulfovibrio_vulgaris_subsp_vulgaris_str_Hildenborough’:2.47E-4):0.096556,’Desulfovibrio_desulfuricans_subsp_desulfuricans_str_G20′:0.130391):0.041935,’Lawsonia_intracellularis_PHE_MN1_00′:0.194671):0.108731,(‘GEBA_Desulfohalobium_retbaense’:0.206905,’GEBA_Desulfomicrobium_baculatum’:0.21572):0.039647):0.204436,’Desulfotalea_psychrophila_LSv54′:0.368324):0.05185,(‘Desulfococcus_oleovorans_Hxd3′:0.325853,’Syntrophobacter_fumaroxidans_MPOB’:0.267991):0.045811):0.045242,’Syntrophus_aciditrophicus_SB’:0.335728):0.039815):0.036306,(((‘Sorangium_cellulosum_So_ce_56_’:0.340758,’GEBA_Haliangium_ochraceum’:0.326975):0.069132,((‘Anaeromyxobacter_sp_Fw109_5′:0.05281,’Anaeromyxobacter_dehalogenans_2CP_C’:0.046187):0.14976,’Myxococcus_xanthus_DK_1622′:0.199974):0.121235):0.065096,’Bdellovibrio_bacteriovorus_HD100′:0.487944):0.045793):0.04331):0.033782):0.033693,’GEBA_Denitrovibrio_acetiphilus’:0.52279):0.036282,(((((((‘Sulfurimonas_denitrificans_DSM_1251′:0.23347,’Arcobacter_butzleri_RM4018′:0.208618):0.040632,’Sulfurovum_sp_NBC37_1’:0.230443):0.030751,(((((‘Campylobacter_jejuni_subsp_jejuni_NCTC_11168′:0.001263,’Campylobacter_jejuni_RM1221’:9.27E-4):6.61E-4,((‘Campylobacter_jejuni_subsp_doylei_269_97′:0.006051,’Campylobacter_jejuni_subsp_jejuni_81116′:3.16E-4):0.001123,’Campylobacter_jejuni_subsp_jejuni_81_176’:9.26E-4):3.5E-4):0.119606,((‘Campylobacter_curvus_525_92′:0.03145,’Campylobacter_concisus_13826′:0.037212):0.061724,’Campylobacter_fetus_subsp_fetus_82_40′:0.097271):0.023098):0.020182,’Campylobacter_hominis_ATCC_BAA_381′:0.16927):0.075991,’GEBA_Sulfurospirillum_deleyianum’:0.136694):0.07072):0.026032,((((((‘Helicobacter_pylori_HPAG1′:0.003032,’Helicobacter_pylori_26695′:0.004293):9.24E-4,’Helicobacter_pylori_Shi470′:0.004956):0.002035,’Helicobacter_pylori_J99′:0.00625):0.008252,’Helicobacter_acinonychis_str_Sheeba’:0.010673):0.199339,’Helicobacter_hepaticus_ATCC_51449′:0.121696):0.062387,’Wolinella_succinogenes_DSM_1740′:0.107844):0.108511):0.052813,’Nitratiruptor_sp_SB155_2′:0.132215):0.401479,(‘Aquifex_aeolicus_VF5′:0.291771,’Sulfurihydrogenibium_sp_YO3AOP1′:0.270137):0.202377):0.049843,’Elusimicrobium_minutum_Pei191’:0.667875):0.034753):0.023181,(((((((‘GEBA_Dyadobacter_fermentans’:0.115999,’GEBA_Spirosoma_linguale’:0.124662):0.074785,’Cytophaga_hutchinsonii_ATCC_33406′:0.182703):0.054101,’Candidatus_Amoebophilus_asiaticus_5a2′:0.332349):0.041954,((‘GEBA_Pedobacter_heparinus’:0.181859,’GEBA_Chitinophaga_pinensis’:0.30193):0.037197,(((((‘Flavobacterium_psychrophilum_JIP02_86′:0.059237,’Flavobacterium_johnsoniae_UW101′:0.052582):0.072716,’Gramella_forsetii_KT0803′:0.133872):0.034419,’GEBA_Capnocytophaga_ochracea’:0.123672):0.087924,’Candidatus_Sulcia_muelleri_GWSS’:0.665368):0.057908,((((‘Bacteroides_fragilis_YCH46′:0.0,’Bacteroides_fragilis_NCTC_9343′:0.0):0.024113,’Bacteroides_thetaiotaomicron_VPI_5482′:0.02726):0.02733,’Bacteroides_vulgatus_ATCC_8482’:0.051865):0.063951,((‘Porphyromonas_gingivalis_ATCC_33277′:8.58E-4,’Porphyromonas_gingivalis_W83′:0.001013):0.151787,’Parabacteroides_distasonis_ATCC_8503’:0.06466):0.046245):0.18136):0.057273):0.041539):0.194704,(‘GEBA_Rhodothermus_marinus’:0.154571,’Salinibacter_ruber_DSM_13855′:0.312844):0.159944):0.062353,((((‘Chlorobium_tepidum_TLS’:0.032082,’Chlorobaculum_parvum_NCIB_8327′:0.032712):0.058237,(((‘Prosthecochloris_vibrioformis_DSM_265′:0.059493,’Pelodictyon_luteolum_DSM_273′:0.0478):0.035359,’Chlorobium_chlorochromatii_CaD3’:0.105899):0.014378,(‘Chlorobium_limicola_DSM_245′:0.051253,’Chlorobium_phaeobacteroides_DSM_266′:0.062034):0.016101):0.038256):0.042465,’Chlorobium_phaeobacteroides_BS1′:0.115118):0.129491,’Chloroherpeton_thalassium_ATCC_35110’:0.162073):0.253695):0.148638,((((‘Akkermansia_muciniphila_ATCC_BAA_835′:0.396966,’Opitutus_terrae_PB90_1′:0.451463):0.059359,’Methylacidiphilum_infernorum_V4’:0.427955):0.105972,(((((‘Chlamydia_trachomatis_D_UW_3_CX’:0.002254,’Chlamydia_trachomatis_A_HAR_13′:0.002736):0.004197,(‘Chlamydia_trachomatis_434_Bu’:0.0,’Chlamydia_trachomatis_L2b_UCH_1_proctitis’:3.44E-4):0.002396):0.021326,’Chlamydia_muridarum_Nigg’:0.019605):0.087368,(((‘Chlamydophila_abortus_S26_3′:0.030285,’Chlamydophila_caviae_GPIC’:0.023437):0.007338,’Chlamydophila_felis_Fe_C_56′:0.022986):0.042096,((‘Chlamydophila_pneumoniae_AR39′:0.0,’Chlamydophila_pneumoniae_J138’:1.71E-4):1.71E-4,(‘Chlamydophila_pneumoniae_CWL029′:3.42E-4,’Chlamydophila_pneumoniae_TW_183′:3.44E-4):0.0):0.088464):0.034515):0.269463,’Candidatus_Protochlamydia_amoebophila_UWE25’:0.26373):0.294159):0.072779,(‘Rhodopirellula_baltica_SH_1′:0.328464,’GEBA_Planctomyces_limnophilus’:0.334605):0.359784):0.051776):0.041976):0.020277,((((((‘Borrelia_afzelii_PKo’:0.011852,’Borrelia_garinii_PBi’:0.016443):0.006519,’Borrelia_burgdorferi_B31′:0.013496):0.093741,’Borrelia_hermsii_DAH’:0.090331):0.391295,((‘Treponema_pallidum_subsp_pallidum_SS14′:0.0,’Treponema_pallidum_subsp_pallidum_str_Nichols’:0.0):0.267368,’Treponema_denticola_ATCC_35405′:0.151221):0.231493):0.155836,’GEBA_Brachyspira_murdochii’:0.52097):0.062841,(((‘Leptospira_borgpetersenii_serovar_Hardjo_bovis_L550′:2.2E-4,’Leptospira_borgpetersenii_serovar_Hardjo_bovis_JB197’:0.0):0.017229,(‘Leptospira_interrogans_serovar_Lai_str_56601′:1.81E-4,’Leptospira_interrogans_serovar_Copenhageni_str_Fiocruz_L1_130’:8.38E-4):0.020842):0.144917,(‘Leptospira_biflexa_serovar_Patoc_strain_Patoc_1_Paris_’:0.0,’Leptospira_biflexa_serovar_Patoc_strain_Patoc_1_Ames_’:1.71E-4):0.165532):0.395767):0.1053):0.036183,((((((((‘Mycoplasma_hyopneumoniae_J’:0.002353,’Mycoplasma_hyopneumoniae_7448′:0.003734):0.002271,’Mycoplasma_hyopneumoniae_232′:0.003699):0.430983,((((‘Mycoplasma_synoviae_53′:0.261036,’Mycoplasma_agalactiae_PG2′:0.263813):0.099129,’Mycoplasma_pulmonis_UAB_CTIP’:0.278073):0.046282,’Mycoplasma_arthritidis_158L3_1′:0.383762):0.035874,’Mycoplasma_mobile_163K’:0.328229):0.046689):0.262618,(((‘Ureaplasma_parvum_serovar_3_str_ATCC_700970′:1.66E-4,’Ureaplasma_parvum_serovar_3_str_ATCC_27815′:0.0):0.453388,’Mycoplasma_penetrans_HF_2’:0.366678):0.056306,((‘Mycoplasma_pneumoniae_M129′:0.106972,’Mycoplasma_genitalium_G37′:0.117431):0.32371,’Mycoplasma_gallisepticum_R’:0.299834):0.137813):0.242873):0.06613,((‘Mycoplasma_mycoides_subsp_mycoides_SC_str_PG1′:0.018411,’Mycoplasma_capricolum_subsp_capricolum_ATCC_27343′:0.013116):0.146931,’Mesoplasma_florum_L1’:0.163211):0.286922):0.11052,(((‘Onion_yellows_phytoplasma_OY_M’:0.016914,’Aster_yellows_witches_broom_phytoplasma_AYWB’:0.018909):0.202409,’Candidatus_Phytoplasma_mali’:0.277597):0.160881,’Acholeplasma_laidlawii_PG_8A’:0.249163):0.180723):0.122978,(((‘GEBA_Streptobacillus_moniliformis’:0.173665,’GEBA_Leptotrichia_buccalis’:0.08471):0.045217,’GEBA_Sebaldella_termitidis’:0.115851):0.114134,’Fusobacterium_nucleatum_subsp_nucleatum_ATCC_25586′:0.22874):0.263641):0.093021,((((((((‘Desulfitobacterium_hafniense_Y51′:0.199501,’Heliobacterium_modesticaldum_Ice1′:0.161842):0.040662,’Moorella_thermoacetica_ATCC_39073’:0.189602):0.025151,((((‘Desulfotomaculum_reducens_MI_1′:0.141921,’GEBA_Desulfotomaculum_acetoxidans’:0.173079):0.026808,’Pelotomaculum_thermopropionicum_SI’:0.119406):0.031051,’Candidatus_Desulforudis_audaxviator_MP104C’:0.239396):0.041262,’Carboxydothermus_hydrogenoformans_Z_2901′:0.192197):0.033883):0.022877,’Syntrophomonas_wolfei_subsp_wolfei_str_Goettingen’:0.340927):0.029485,’Symbiobacterium_thermophilum_IAM_14863′:0.302816):0.024169,(‘Natranaerobius_thermophilus_JW_NM_WN_LF’:0.328273,’GEBA_Veillonella_parvula’:0.318108):0.03302):0.020798,((((((((((((((‘Bacillus_thuringiensis_serovar_konkukian_str_97_27′:3.38E-4,’Bacillus_thuringiensis_str_Al_Hakam’:1.69E-4):1.69E-4,((‘Bacillus_anthracis_str_Sterne’:0.0,’Bacillus_anthracis_str_Ames_Ancestor_’:0.0):0.0,’Bacillus_anthracis_str_Ames’:0.0):0.001183):6.51E-4,’Bacillus_cereus_E33L’:5.32E-4):0.001131,(‘Bacillus_cereus_ATCC_14579′:0.002251,’Bacillus_weihenstephanensis_KBAB4′:0.013987):0.004118):0.001515,’Bacillus_cereus_ATCC_10987′:2.57E-4):0.020967,’Bacillus_cereus_subsp_cytotoxis_NVH_391_98’:0.013733):0.085584,(((‘Bacillus_subtilis_subsp_subtilis_str_168′:0.015012,’Bacillus_amyloliquefaciens_FZB42′:0.015183):0.020109,’Bacillus_pumilus_SAFR_032′:0.040049):0.011797,’Bacillus_licheniformis_ATCC_14580’:0.023456):0.065389):0.021459,(‘Geobacillus_thermodenitrificans_NG80_2′:0.011367,’Geobacillus_kaustophilus_HTA426’:0.01873):0.094956):0.020796,((((((((‘Staphylococcus_aureus_subsp_aureus_Mu3′:0.0,’Staphylococcus_aureus_subsp_aureus_Mu50′:1.67E-4):1.67E-4,’Staphylococcus_aureus_subsp_aureus_N315’:1.67E-4):0.0,(‘Staphylococcus_aureus_subsp_aureus_JH9′:0.001001,’Staphylococcus_aureus_subsp_aureus_JH1’:0.0):3.34E-4):5.16E-4,(((‘Staphylococcus_aureus_subsp_aureus_MRSA252′:6.67E-4,’Staphylococcus_aureus_RF122’:3.35E-4):0.0,(((‘Staphylococcus_aureus_subsp_aureus_NCTC_8325′:6.77E-4,’Staphylococcus_aureus_subsp_aureus_USA300_TCH1516′:0.0):0.0,’Staphylococcus_aureus_subsp_aureus_USA300’:0.0):0.0,(‘Staphylococcus_aureus_subsp_aureus_COL’:6.68E-4,’Staphylococcus_aureus_subsp_aureus_str_Newman’:3.33E-4):3.34E-4):1.67E-4):0.0,(‘Staphylococcus_aureus_subsp_aureus_MSSA476′:0.0,’Staphylococcus_aureus_subsp_aureus_MW2’:0.0):3.34E-4):1.51E-4):0.032187,((‘Staphylococcus_epidermidis_ATCC_12228′:3.35E-4,’Staphylococcus_epidermidis_RP62A’:1.66E-4):0.023543,’Staphylococcus_haemolyticus_JCSC1435′:0.025193):0.012868):0.014679,’Staphylococcus_saprophyticus_subsp_saprophyticus_ATCC_15305′:0.045225):0.208482,((((((((((‘Streptococcus_pyogenes_SSI_1′:1.69E-4,’Streptococcus_pyogenes_MGAS315’:3.35E-4):3.86E-4,(((((‘Streptococcus_pyogenes_M1_GAS’:3.34E-4,’Streptococcus_pyogenes_MGAS5005′:1.67E-4):3.34E-4,(((‘Streptococcus_pyogenes_MGAS8232′:6.78E-4,’Streptococcus_pyogenes_MGAS10750′:8.62E-4):1.67E-4,’Streptococcus_pyogenes_str_Manfredo’:5.01E-4):3.34E-4,’Streptococcus_pyogenes_MGAS10394′:5.01E-4):0.0):1.67E-4,(‘Streptococcus_pyogenes_MGAS9429′:5.01E-4,’Streptococcus_pyogenes_MGAS2096′:0.0):5.01E-4):0.0,’Streptococcus_pyogenes_MGAS10270′:5.09E-4):3.34E-4,’Streptococcus_pyogenes_MGAS6180’:5.01E-4):2.83E-4):0.032705,((‘Streptococcus_agalactiae_2603V_R’:5.02E-4,’Streptococcus_agalactiae_A909′:0.0):5.02E-4,’Streptococcus_agalactiae_NEM316′:0.0):0.032531):0.009744,((‘Streptococcus_thermophilus_LMG_18311′:4.96E-4,’Streptococcus_thermophilus_CNRZ1066′:6.75E-4):0.001002,’Streptococcus_thermophilus_LMD_9′:8.46E-4):0.042346):0.010989,’Streptococcus_mutans_UA159’:0.053241):0.014154,((‘Streptococcus_suis_98HAH33′:1.41E-4,’Streptococcus_suis_05ZYH33’:0.003069):0.045053,((((‘Streptococcus_pneumoniae_D39′:0.0,’Streptococcus_pneumoniae_R6’:0.0):0.001089,(‘Streptococcus_pneumoniae_TIGR4′:7.57E-4,’Streptococcus_pneumoniae_CGSP14′:0.001274):5.04E-4):1.79E-4,’Streptococcus_pneumoniae_Hungary19A_6’:5.78E-4):0.025464,(‘Streptococcus_gordonii_str_Challis_substr_CH1′:0.013866,’Streptococcus_sanguinis_SK36’:0.013168):0.014048):0.01685):0.015608):0.079689,((‘Lactococcus_lactis_subsp_cremoris_SK11′:0.001565,’Lactococcus_lactis_subsp_cremoris_MG1363′:8.56E-4):0.005333,’Lactococcus_lactis_subsp_lactis_Il1403′:0.005458):0.157584):0.124114,’Enterococcus_faecalis_V583’:0.096988):0.030082,(((((‘Lactobacillus_acidophilus_NCFM’:0.017053,’Lactobacillus_helveticus_DPC_4571′:0.025039):0.047802,(‘Lactobacillus_gasseri_ATCC_33323′:0.008251,’Lactobacillus_johnsonii_NCC_533’:0.00468):0.081515):0.035749,(‘Lactobacillus_delbrueckii_subsp_bulgaricus_ATCC_BAA_365′:0.001075,’Lactobacillus_delbrueckii_subsp_bulgaricus_ATCC_11842’:9.47E-4):0.095385):0.189762,((‘Lactobacillus_casei_BL23′:1.67E-4,’Lactobacillus_casei_ATCC_334′:1.71E-4):0.13496,’Lactobacillus_sakei_subsp_sakei_23K’:0.116042):0.029638):0.037981,(((((‘Lactobacillus_brevis_ATCC_367′:0.113171,’Lactobacillus_plantarum_WCFS1′:0.09989):0.023598,’Pediococcus_pentosaceus_ATCC_25745’:0.140539):0.018317,(‘Lactobacillus_fermentum_IFO_3956′:0.075472,’Lactobacillus_reuteri_F275’:0.062813):0.114981):0.022679,((‘Leuconostoc_mesenteroides_subsp_mesenteroides_ATCC_8293′:0.031056,’Leuconostoc_citreum_KM20′:0.035589):0.124121,’Oenococcus_oeni_PSU_1′:0.234536):0.131959):0.021585,’Lactobacillus_salivarius_UCC118’:0.124879):0.034651):0.077386):0.111668,(((‘Listeria_monocytogenes_EGD_e’:0.00208,’Listeria_monocytogenes_str_4b_F2365′:7.75E-4):0.001511,’Listeria_welshimeri_serovar_6b_str_SLCC5334′:0.003974):0.001047,’Listeria_innocua_Clip11262′:0.001031):0.107265):0.033808):0.044404,’Lysinibacillus_sphaericus_C3_41′:0.152339):0.036331):0.021337,’Oceanobacillus_iheyensis_HTE831′:0.18896):0.019902,(‘Bacillus_clausii_KSM_K16′:0.093196,’Bacillus_halodurans_C_125′:0.066964):0.053506):0.027554,’Exiguobacterium_sibiricum_255_15′:0.211271):0.115095,’GEBA_Alicyclobacillus_acidocaldarius’:0.223192):0.06559,(((((‘Clostridium_acetobutylicum_ATCC_824′:0.123954,’Clostridium_novyi_NT’:0.114099):0.023517,((((‘Clostridium_perfringens_ATCC_13124′:5.13E-4,’Clostridium_perfringens_str_13′:0.001376):4.86E-4,’Clostridium_perfringens_SM101’:0.00102):0.09186,((‘Clostridium_botulinum_E3_str_Alaska_E43′:0.003782,’Clostridium_botulinum_B_str_Eklund_17B’:0.003812):0.055674,’Clostridium_beijerinckii_NCIMB_8052′:0.067314):0.054513):0.057627,((((((‘Clostridium_botulinum_A_str_ATCC_3502′:0.0,’Clostridium_botulinum_A_str_ATCC_19397′:0.0):0.0,’Clostridium_botulinum_A_str_Hall’:1.67E-4):0.001673,(‘Clostridium_botulinum_F_str_Langeland’:0.001211,’Clostridium_botulinum_B1_str_Okra’:0.003486):0.001309):0.001791,’Clostridium_botulinum_A3_str_Loch_Maree’:0.007218):0.098641,’Clostridium_kluyveri_DSM_555′:0.121865):0.020585,’Clostridium_tetani_E88′:0.115493):0.020236):0.012084):0.136573,’Clostridium_phytofermentans_ISDg’:0.312208):0.036695,((‘GEBA_Anaerococcus_prevotii’:0.28787,’Finegoldia_magna_ATCC_29328′:0.211317):0.173769,((‘Alkaliphilus_metalliredigens_QYMF’:0.12868,’Alkaliphilus_oremlandii_OhILAs’:0.107044):0.082634,’Clostridium_difficile_630′:0.211875):0.045791):0.027696):0.039355,((((‘Thermoanaerobacter_pseudethanolicus_ATCC_33223′:0.004904,’Thermoanaerobacter_sp_X514′:0.004594):0.034053,’Thermoanaerobacter_tengcongensis_MB4′:0.041493):0.149566,’Caldicellulosiruptor_saccharolyticus_DSM_8903′:0.23409):0.036154,’Clostridium_thermocellum_ATCC_27405’:0.176618):0.036512):0.061826):0.019474):0.069313,((((((((((((((‘GEBA_Sanguibacter_keddieii’:0.064063,’GEBA_Jonesia_denitrificans’:0.099947):0.030002,’GEBA_Xylanimonas_cellulosilytica’:0.094672):0.01975,’GEBA_Cellulomonas_flavigena’:0.082405):0.035617,’GEBA_Beutenbergia_cavernae’:0.115349):0.051352,((‘GEBA_Brachybacterium_faecium’:0.196353,’GEBA_Kytococcus_sedentarius’:0.168724):0.036428,((((‘Arthrobacter_aurescens_TC1′:0.020568,’Arthrobacter_sp_FB24′:0.028887):0.039891,’Renibacterium_salmoninarum_ATCC_33209′:0.068094):0.050028,’Kocuria_rhizophila_DC2201’:0.130484):0.065937,((‘Clavibacter_michiganensis_subsp_michiganensis_NCPPB_382′:0.003996,’Clavibacter_michiganensis_subsp_sepedonicus’:0.003983):0.084663,’Leifsonia_xyli_subsp_xyli_str_CTCB07′:0.089834):0.133992):0.04523):0.013623):0.017241,’Kineococcus_radiotolerans_SRS30216′:0.154573):0.033958,(((((((‘GEBA_Nocardiopsis_dassonvillei’:0.093463,’Thermobifida_fusca_YX’:0.072507):0.074571,’GEBA_Thermomonospora_curvata’:0.099223):0.020272,(‘GEBA_Thermobispora_bispora’:0.072121,’GEBA_Streptosporangium_roseum’:0.083245):0.048392):0.038128,’Acidothermus_cellulolyticus_11B’:0.153492):0.027393,(((‘Frankia_sp_CcI3′:0.019914,’Frankia_alni_ACN14a’:0.017049):0.027318,’Frankia_sp_EAN1pec’:0.039855):0.137522,((((((((((‘Mycobacterium_gilvum_PYR_GCK’:0.025373,’Mycobacterium_vanbaalenii_PYR_1′:0.014398):0.020335,((‘Mycobacterium_sp_KMS’:0.0,’Mycobacterium_sp_MCS’:0.0):2.54E-4,’Mycobacterium_sp_JLS’:4.38E-4):0.029295):0.012695,’Mycobacterium_smegmatis_str_MC2_155′:0.027998):0.015961,((((((((‘Mycobacterium_bovis_AF2122_97′:0.0,’Mycobacterium_bovis_BCG_str_Pasteur_1173P2′:3.48E-4):5.22E-4,’Mycobacterium_tuberculosis_F11′:3.48E-4):0.0,’Mycobacterium_tuberculosis_H37Ra’:0.0):0.0,’Mycobacterium_tuberculosis_H37Rv’:0.0):1.74E-4,’Mycobacterium_tuberculosis_CDC1551′:3.48E-4):0.028503,(‘Mycobacterium_marinum_M’:2.83E-4,’Mycobacterium_ulcerans_Agy99′:0.005107):0.026919):0.008581,’Mycobacterium_leprae_TN’:0.050438):0.008421,(‘Mycobacterium_avium_104′:1.71E-4,’Mycobacterium_avium_subsp_paratuberculosis_K_10′:0.001878):0.024032):0.041807):0.021744,’Mycobacterium_abscessus’:0.054563):0.05338,((‘GEBA_Gordonia_bronchialis’:0.080084,’GEBA_Tsukamurella_paurometabola’:0.095182):0.023186,(‘Nocardia_farcinica_IFM_10152′:0.066164,’Rhodococcus_sp_RHA1’:0.059553):0.015357):0.01873):0.081897,((‘GEBA_Saccharomonospora_viridis’:0.098036,’Saccharopolyspora_erythraea_NRRL_2338′:0.100342):0.021978,’GEBA_Actinosynnema_mirum’:0.097173):0.027185):0.031045,’GEBA_Nakamurella_multipartita’:0.151197):0.032028,’GEBA_Geodermatophilus_obscurus’:0.140115):0.027068,((‘Salinispora_arenicola_CNS_205′:0.012069,’Salinispora_tropica_CNB_440′:0.010019):0.097618,’GEBA_Stackebrandtia_nassauensis’:0.170224):0.084563):0.033468):0.03169):0.026445,(((‘Streptomyces_avermitilis_MA_4680′:0.027139,’Streptomyces_coelicolor_A3_2_’:0.027351):0.021484,’Streptomyces_griseus_subsp_griseus_NBRC_13350′:0.037328):0.090414,’GEBA_Catenulispora_acidiphila’:0.137426):0.05113):0.0286,((‘GEBA_Kribbella_flavida’:0.118628,’Nocardioides_sp_JS614′:0.133604):0.026332,’Propionibacterium_acnes_KPA171202′:0.242956):0.044904):0.019561):0.056195,((((‘Corynebacterium_glutamicum_R’:4.77E-4,’Corynebacterium_glutamicum_ATCC_13032′:0.001073):0.033614,’Corynebacterium_efficiens_YS_314′:0.038791):0.051561,’Corynebacterium_diphtheriae_NCTC_13129′:0.068116):0.044622,(‘Corynebacterium_urealyticum_DSM_7109′:0.065695,’Corynebacterium_jeikeium_K411’:0.054499):0.056416):0.221144):0.045301,((‘Bifidobacterium_longum_DJO10A’:1.88E-4,’Bifidobacterium_longum_NCC2705′:3.39E-4):0.040933,’Bifidobacterium_adolescentis_ATCC_15703′:0.037617):0.303161):0.039753,(‘Tropheryma_whipplei_TW08_27′:8.95E-4,’Tropheryma_whipplei_str_Twist’:6.79E-4):0.478476):0.120846,’GEBA_Acidimicrobium_ferrooxidans’:0.43829):0.090081,(((‘GEBA_Cryptobacterium_curtum’:0.142154,’GEBA_Eggerthella_lenta’:0.088587):0.047811,’GEBA_Slackia_heliotrinireducens’:0.117231):0.104268,’GEBA_Atopobium_parvulum’:0.269623):0.198082):0.036936,(‘GEBA_Conexibacter_woesei’:0.383152,’Rubrobacter_xylanophilus_DSM_9941′:0.357989):0.109409):0.087352,(((((((((‘Synechococcus_sp_WH_7803′:0.023543,’Synechococcus_sp_CC9311’:0.044194):0.014221,((‘Synechococcus_sp_CC9605′:0.022222,’Synechococcus_sp_WH_8102′:0.025798):0.006423,’Synechococcus_sp_CC9902’:0.032326):0.022152):0.016325,(((‘Prochlorococcus_marinus_subsp_marinus_str_CCMP1375′:0.058394,’Prochlorococcus_marinus_str_MIT_9211’:0.053438):0.019947,((‘Prochlorococcus_marinus_str_NATL2A’:0.00276,’Prochlorococcus_marinus_str_NATL1A’:0.002109):0.070906,((‘Prochlorococcus_marinus_str_MIT_9515′:0.013908,’Prochlorococcus_marinus_subsp_pastoris_str_CCMP1986’:0.016702):0.023743,(((‘Prochlorococcus_marinus_str_AS9601′:0.005031,’Prochlorococcus_marinus_str_MIT_9301′:0.005416):0.003975,’Prochlorococcus_marinus_str_MIT_9215′:0.011185):0.004908,’Prochlorococcus_marinus_str_MIT_9312’:0.010415):0.020455):0.117983):0.022565):0.040798,(‘Prochlorococcus_marinus_str_MIT_9313′:0.00429,’Prochlorococcus_marinus_str_MIT_9303′:0.003453):0.043557):0.012995):0.046129,’Synechococcus_sp_RCC307’:0.067465):0.195828,(‘Synechococcus_elongatus_PCC_7942′:3.21E-4,’Synechococcus_elongatus_PCC_6301’:0.00142):0.098101):0.061386,(((((‘Nostoc_sp_PCC_7120′:0.004827,’Anabaena_variabilis_ATCC_29413′:0.003501):0.036361,’Nostoc_punctiforme_PCC_73102′:0.042386):0.086848,’Trichodesmium_erythraeum_IMS101’:0.151445):0.024405,(((‘Cyanothece_sp_ATCC_51142′:0.088208,’Microcystis_aeruginosa_NIES_843′:0.108904):0.018975,’Synechocystis_sp_PCC_6803′:0.111161):0.027275,’Synechococcus_sp_PCC_7002’:0.127272):0.053965):0.023778,(‘Acaryochloris_marina_MBIC11017′:0.127345,’Thermosynechococcus_elongatus_BP_1’:0.138774):0.035212):0.017944):0.063385,(‘Synechococcus_sp_JA_3_3Ab’:0.023735,’Synechococcus_sp_JA_2_3B_a_2_13_’:0.025648):0.172906):0.056598,’Gloeobacter_violaceus_PCC_7421′:0.211642):0.301851,(((((‘Roseiflexus_castenholzii_DSM_13941′:0.02822,’Roseiflexus_sp_RS_1′:0.017828):0.118476,’Chloroflexus_aurantiacus_J_10_fl’:0.154341):0.066504,’Herpetosiphon_aurantiacus_ATCC_23779′:0.219302):0.124086,(‘GEBA_Sphaerobacter_thermophilus’:0.242004,’GEBA_Thermobaculum_terrenum’:0.272557):0.055029):0.055648,((‘Dehalococcoides_sp_CBDB1′:5.54E-4,’Dehalococcoides_sp_BAV1′:8.51E-4):0.017643,’Dehalococcoides_ethenogenes_195’:0.014048):0.50406):0.083393):0.032459):0.039409):0.035188):0.022061):0.039665):0.054226,(((‘Thermosipho_melanesiensis_BI429′:0.127803,’Fervidobacterium_nodosum_Rt17_B1’:0.188887):0.074834,(((‘Thermotoga_maritima_MSB8′:0.007577,’Thermotoga_sp_RQ2′:5.3E-4):0.005057,’Thermotoga_petrophila_RKU_1′:0.006656):0.159923,’Thermotoga_lettingae_TMO’:0.230211):0.037393):0.080613,’Petrotoga_mobilis_SJ95′:0.362128):0.246177):0.287154,((‘Thermus_thermophilus_HB8′:0.001498,’Thermus_thermophilus_HB27’:0.001224):0.144321,(‘GEBA_Meiothermus_ruber’:0.114423,’GEBA_Meiothermus_silvanus’:0.077947):0.086289):0.12529):0.1212895,(‘Deinococcus_geothermalis_DSM_11300′:0.043888,’Deinococcus_radiodurans_R1’:0.07314):0.1212895);
END;