There is a wonderful paper that has just appeared in PLoS Genetics I want to call people’s attention to: PLoS Genetics: Emergence and Modular Evolution of a Novel Motility Machinery in Bacteria
In the paper, researchers from CNRS and Aix-Marseille in France used some nice comparative and evolutionary genomics analyses along with experimental work to characterize the function and evolution of gliding motility in bacteria.
Their summary of their work:
Motility over solid surfaces (gliding) is an important bacterial mechanism that allows complex social behaviours and pathogenesis. Conflicting models have been suggested to explain this locomotion in the deltaproteobacterium Myxococcus xanthus: propulsion by polymer secretion at the rear of the cells as opposed to energized nano-machines distributed along the cell body. However, in absence of characterized molecular machinery, the exact mechanism of gliding could not be resolved despite several decades of research. In this study, using a combination of experimental and computational approaches, we showed for the first time that the motility machinery is composed of large macromolecular assemblies periodically distributed along the cell envelope. Furthermore, the data suggest that the motility machinery derived from an ancient gene cluster also found in several non-gliding bacterial lineages. Intriguingly, we find that most of the components of the gliding machinery are closely related to a sporulation system, suggesting unsuspected links between these two apparently distinct biological processes. Our findings now pave the way for the first molecular studies of a long mysterious motility mechanism.
Basically, they started with some genetic and functional studies in Myxococcus xanthus. They analyzed these in the context of the genome sequence (note – I was a co-author on the original genome paper). And then they did some extensive comparative and evolutionary analysis of these genes, producing some wonderful figures along the way such as:
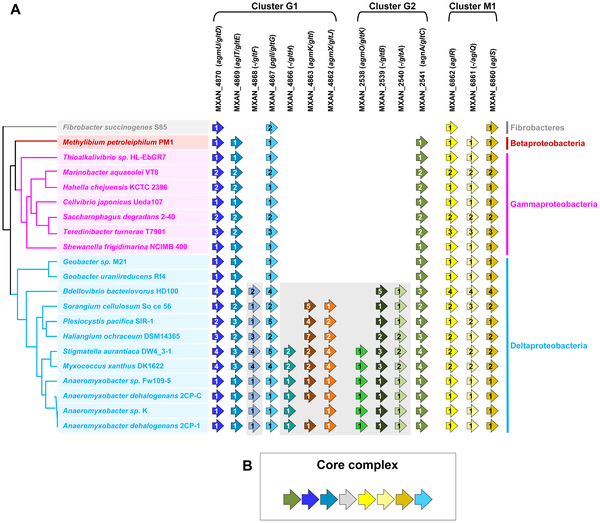 |
| Figure 2. Taxonomic distribution of the closest homologues of the 14 genes composing the G1, G2, and M1 clusters, and genetic organization of the core complex. (A) For a given gene, the number of homologues in the corresponding genome is indicated by the numbers within arrows. The relationships between the species carrying the different homologues of the genes are indicated by the phylogeny on the left. Based on their taxonomic distribution, the 14 genes can be divided into Group A (grey background) and Group B (white background). (B) In all non Deltaproteobacteria and in Geobacter, the Group B genes clustered in a single genomic region. doi:10.1371/journal.pgen.1002268.g002 |
Based on their analysis they then came up with some hypotheses as to which genes were involved in key parts of gliding motility and what their biochemical functions were and they then went and confirmed this with experiments. I am not going to go into detail on the functional work they did but you can read their paper for more details.
They wrapped up their paper by proposing an model for the evolutionary history of gliding motility. I am not sure I buy all components of their model since our sampling of genomes right now is still very poor, but they have a pretty detailed theory captured in part in this figure:
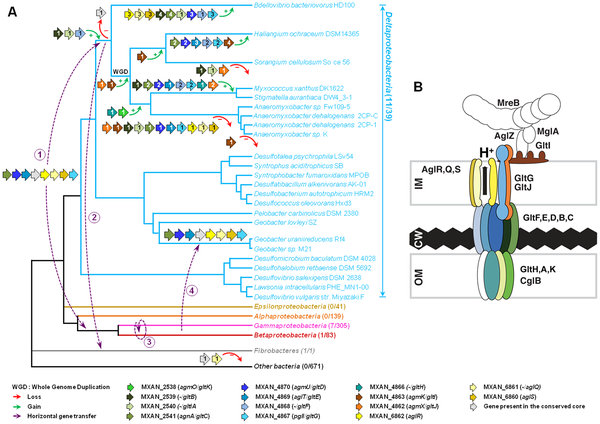 |
| Figure 8. Evolution and structure of the Myxococcus gliding motility machinery. A) Evolutionary scenario describing the emergence and evolution of the gliding motility machinery in M. xanthus. The relationships between organisms carrying close homologues of the 14 genes encoding putative components of the gliding machinery in M. xanthus are represented by the phylogeny. Green and red arrows respectively indicate gene acquisition and gene loss. The number of gene copies that were acquired or lost is indicated within arrows. The purple dotted arrows represent horizontal gene transfer events of one or several components. WGD marks the putative whole genome duplication event that occurred in the ancestor of Myxococcales. For each gene, locus_tag, former (agm/agl/agn) and new (glt and agl) names are provided. The number of complete genomes that contain homologues of glt and agl genes compared to the total number of complete genomes available at the beginning of this study are indicated in brackets. (B) The Myxococcus gliding machinery. The diagram compiles data from this work and published literature. Components were added based on bioinformatic predictions, mutagenesis, interaction and localization studies. Exhaustive information is not available for all proteins and thus the diagram largely is subject to modifications once more data will be available. Known interactions within the complex from experimental evidence are AglR-GltG, AglZ-MglA and interactions within the AglRQS molecular motor [13], [15]. For clarity, the proteins were colour-coded as in the rest of the manuscript |
Anyway – I don’t have much time right now to provide more detail on the paper. But it is definitely worth checking out.